
Stuart Kornfeld, MD
David C. and Betty Farrell Distinguished Professor of Medicine
- Phone: 314-362-8803
- Fax: 314-362-8826
- Email: skornfel@nospam.wustl.edu
Address:
- Hematology Division
Mail Stop 8125-0020-08, Washington University, 660 South Euclid Avenue, St. Louis, MO 63110 (mail) - Room 8826 Clinical Sciences Research Building (office)
- Room 8828 Clinical Sciences Research Building (lab)
Clinical Interest: General hematology
Research Interest: Intracellular protein trafficking; Lysosomes; Glycoproteins
Research: We are studying the biochemical and molecular basis for protein trafficking in mammalian cells, focusing on the biogenesis of lysosomes. The targeting of lysosomal enzymes from their site of synthesis in the rough endoplasmic reticulum to their final destination in lysosomes is a multistep process that utilizes a series of recognition signals that must be deciphered by cellular components that mediate the sorting, packaging and transport of the lysosomal enzymes. A key step in this pathway is the selective phosphorylation of mannose residues on the lysosomal enzymes. These residues allow binding to Man-6-P receptors in the Golgi and transport to lysosomes via clathrin-coated vesicles.
The goal of our work is to understand this complex sorting system at the molecular level. Currently we are studying how the enzyme that mediates mannose phosphorylation recognizes lysosomal enzymes as specific substrates and how the Man-6-P receptors bind their ligands, enter clathrin-coated pits, and traffic between various cellular compartments. Recently we discovered that a newly described family of coat proteins termed GGAs bind the acidic cluster/dileucine lysosomal sorting motif on the cytoplasmic tails of the Man-6-P receptors. Our findings indicate that the GGAs function as connectors to transfer the Man-6-P receptors to AP-1 clathrin-coated vesicles.
We have disrupted the genes in mice that encode the two subunits of the enzyme (N-acetylglucosamine-1-phosphotransferase) that generates the Man-6-P recognition marker on lysosomal enzymes as well as two of the three genes that encode the GGAs. These mice are allowing us to dissect the role of each component of this targeting pathway in the intact animal.
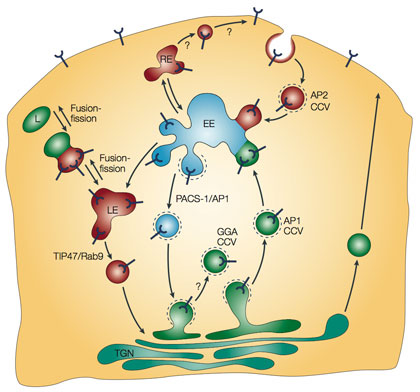
Mannose 6-phosphate receptors (MPRs) bind to their cargo (acid hydrolases) in the trans-Golgi network (TGN), and are packaged into transport carriers that deliver the receptor with its bound ligand to early endosomes (EE). The low pH within the endosomes facilitates the dissociation of the acid hydrolases from the MPRs. Dynamic fusion/fission between the late endosomal and lysosomal compartments results in selective delivery of the hydrolases to the lysosome (L) (the ‘kiss-and-run’ theory77). TIP47/Rab9 prevent the MPRs from reaching the lysosomes, in which they would otherwise be degraded. The return pathway from the early endosomal compartment to the Golgi is probably mediated by PACS-1-assisted packaging into AP1-containing clathrin-coated vesicles (CCVs), whereas that from the late endosomal (LE) compartments is mediated by TIP47 and Rab9. Some of the MPRs go to the cell surface either from early or late endosomes through the recycling endosome (RE), or from proximal TGN cisternae as a consequence of mis-sorting. The cell-surface receptors are internalized in AP2 CCVs and delivered back to the endosomes.
From: Ghosh P, Dahms NM, Kornfeld S
Mannose 6-phosphate receptors: new twists in the tale.
Nat Rev Mol Cell Biol 2003 Mar;4(3):202-12
Biosketch
Education
- 1962: MD, Washington University School of Medicine, St. Louis, MO
- 1958: AB, Dartmouth College, Hanover, NH
Post-Graduate Training
- 1966-1965: Assistant Resident in Ward Medicine, Barnes Hospital, St. Louis, MO
- 1965-1963: Research Associate, National Institute of Arthritis and Metabolic Diseases, NIH
- 1963-1962: Intern in Ward Medicine, Barnes Hospital, St. Louis, MO
Academic Positions & Employment
- 2000-present: David C. and Betty Farrell Distinguished Professor of Medicine
- 1972-present: Professor of Medicine, Washington University School of Medicine
- 2009-2008: Director, Division of Hematology, Washington University School of Medicine
- 2008-1993: Co-Director, Division of Hematology, Washington University School of Medicine
- 1992-1976: Co-Director, Division of Hematology-Oncology, Washington University School of Medicine
- 1976-1973: Director, Division of Oncology, Washington University School of Medicine
- 1972-1970: Associate Professor of Medicine, Washington University School of Medicine
- 1976-1968: Assistant Professor of Biochemistry, Washington University School of Medicine
- 1970-1967: Assistant Professor of Medicine, Washington University School of Medicine
- 1967-1966: Instructor in Medicine, Washington University School of Medicine, St. Louis, MO
Appointments & Committees
- 2018-present: Co-Founder, M6P Therapeutics
- 2019-2000: Co-Director, Physician Scientist Training Program, Washington University School of Medicine
- 1997-1991: Director, Medical Scientist Training Program, Washington University School of Medicine
Honors & Awards
- 2022: Roscoe O. Brady Award, WORLDSymposium (WORLD is an acronym for We’re Organizing Research on Lysosomal Diseases)
- 2021: Honorary Doctor of Science, Washington University
- 2018: Lifetime Achievement Award, St. Louis Academy of Science
- 2017: Lifetime Achievement Award, International Society for Alpha-Mannosidosis and Related Diseases
- 2012: Herbert Tabor/Journal of Biological Chemistry Lectureship, American Society for Biochemistry and Molecular Biology
- 2010: E. B. Wilson Medal, American Society for Cell Biology (with James Rothman and Randy Schekman)
- 2010: Distinguished Faculty Award, Washington University
- 2010: Kober Medal, Association of American Physicians
- 2002: Second Century Award, Washington University
- 2002: UCSD/Nature Medicine “Mentorship Award”
- 2002: Gerty & Carl Cori Faculty Recognition Award, Washington University
- 1999: Karl Meyer Award, Society for Glycobiology
- 1992: E. Donnall Thomas Lectureship and Prize
- 1991: Passano Award (with William Sly)
- 1989: Jubilee Lecturer and Harden Medallist, The Biochemical Society
- 1987: Alumni/Faculty Award, Washington University School of Medicine
- 1984: American Academy of Arts and Sciences
- 1983: National Academy of Medicine
- 1982: National Academy of Sciences
- 1976-1971: Research Career Development Award, National Institutes of Health
- 1971-1966: American Cancer Society Faculty Research Associate
- 1962: Bordon Award for Outstanding Undergraduate Research
- 1959: Biochemistry Award, Washington University School of Medicine
Editorial Responsibilities
- 2007-1997: Consulting Editor, Journal of Clinical Investigation
- 2004-1992: Editorial Board, Molecular Biology of the Cell
- 2003-1998: Editorial Board, Journal of Biological Chemistry
- 1996-1972: Editorial Board, Archives of Biochemistry and Biophysics
- 1991-1988: Editorial Board, Journal of Cell Biology
- 1991-1985: Editorial Board, Proceedings of the National Academy of Sciences
- 1987-1982: Associate Editor, Journal of Biological Chemistry
- 1982-1981: Editor, Journal of Clinical Investigation
- 1981-1977: Associate Editor, Journal of Clinical Investigation
- 1981-1976: Editorial Board, Journal of Biological Chemistry
Professional Societies
- American Society for Clinical Investigation
- American Society of Biological Chemists
- American Society of Hematolgy
- Association of American Physicians
- Foreign Member, Finnish Society of Sciences and Letters
Board Memberships
- 1997-present: Member, Board of Directors, Edward Mallinckrodt, Jr. Foundation
- 2012-1998: Albert Lasker Medical Research Awards Jury
- 2000-1995: Medical Advisory Board, Howard Hughes Medical Institute
- 1998-1997: President, Association of American Physicians
- 1997-1991: Councillor, Associaton of American Physicians
- 1995-1987: Member, Board of Scientific Advisors, Jane Coffin Childs Memorial Fund for Medical Research
- 1994-1986: Member, Scientific Review Board, Howard Hughes Medical Institute
- 1991-1986: Secretary, Association of American Physicians
- 1987-1983: Member, NIADDK Board of Scientific Counselors
- 1977-1974: Member, NIH Cell Biology Study Section
- 1975-1972: Councillor, American Society for Clinical Investigation